08.04.2023|
|
0 Comment
Конспект перелетные птицы в старшей группе: конспект занятия в старшей группе на тему “Перелётный птицы” | План-конспект занятия по окружающему миру (старшая группа):
Конспект занятия по развитию речи «Перелетные птицы» (Старшая группа)
муниципальное бюджетное дошкольное образовательное учреждение
«Детский сад № 198» комбинированного вида
Конспект занятия по развитию речи
«Перелетные птицы»
(Старшая группа)
Составила и провела:
Суевалова Кристина Александровна
Новокузнецк,
Программное содержание:развитие словаря: активизировать название перелетных птиц. Обобщить понятие «перелетные птицы», учить понимать детей образный смысл загадок, формировать грамматический строй речи;
Связанная речь: продолжать учить детей полно и точно отвечать на вопрос, строя предложения из 4-6 слов, развивать навыки составления рассказа по опорным схемам (рисункам).
Оборудование:опорные схемы, фотографии перелетных птиц, мяч.
Ход занятия:
– Ребята сейчас вы поочереди будите называть признаки весны и передавать мяч соседу. Свое предложение начинайте со слов “Весной …..”
Свое предложение начинайте со слов “Весной …..”
– Сегодня мы продолжим разговор о перелетных птицах. Вы уже очень многое знаете о них. Попробуйте отгадать загадки о перелетных птицах.
Черный, проворный,
Кричит “Крак”,
Червякам враг (грач).
– Кто это? И как вы догадались?
– Это грач. Он кричит “Крак!” и любит червяков.
На шесте дворец.
Во дворе певец.
А зовут его… (скворец).
Кто на елке, на суку счет ведет: “Ку-ку. Ку-ку”? (кукушка).
Хочет— прямо полетит,
Хочет — в воздухе висит,
Камнем падает с высот.
И в полях поет, поет (жаворонок).
Прилетает к нам с теплом,
Путь, проделав длинный,
Лепит домик под окном
Из травы и глины (ласточка).
Бродит по болотам грязным,
Ловит в них лягушек,
Бестолковых попрыгушек (цапля, журавль).
Игра “Собери картинку”. [Развитие зрительного гнозиса и конструктивного праксиса.]
Воспитатель раздает детям конверты с разрезными картинками, на которых изображены перелетные птицы.
– Достаньте разрезанные на части картинки и попробуйте собрать их.
Детям, затрудняющимся в выполнении, логопед дает зрительную опору в виде целостной картинки.
– Расскажите, чьи изображения вы сложили.
– Это грач.
– Правильно. Сравни свое изображение с целой картинкой.
– Молодцы. Вы выполнили очень сложное задание. Сложите разрезанные картинки в конверты, а целые положите поближе.- Назови части тела птицы, особенности ее внешнего вида: какие у нее крылья, клюв, лапы? Если ребенок говорит «длинный клюв», воспитатель «значит мы его можем назвать длинноклювым», «черные крылья» – «чернокрылый» и т.д.
Это скворец. Какой у него хвост? (Короткий). Значит скворец какой? – Короткохвостый. Это ласточка, у нее длинный хвост. Значит какая ласточка? – длиннохвостая. Это грач, у него острый клюв. Какой грач? – Остроклювый. У него чёрное крыло? – Чернокрылый.
Все движения разминки повторяем без запинки!
Эй! Попрыгали на месте. Эх! Руками машем вместе.
Эхе – хе! Прогнули спинки, Посмотрели на ботинки.
Эге – ге! Нагнулись ниже, Наклонились к полу ближе.
Повертись на месте ловко. В этом нам нужна сноровка.
Что, понравилось, дружок? Завтра будет вновь урок!
Дидактическое упражнение «Что могут делать птицы?»
Дети, давайте вспомним, что могут делать птицы.
Ходят, клюют, взлетают, летают, выводят, прыгают, вьют, кормят, ухаживают, высиживают, поют.
Игра с мячом “У кого кто”. [Совершенствование грамматического строя речи (образование сущ. с суффиксами –ат, -ят).]
Воспитатель приглашает детей на ковер.
– Давайте поиграем в игру с мячом “У кого кто”. Я буду бросать “звучащий” мячик каждому из вас и называть взрослую птицу. Вы будите ловить мяч, называть птенцов этой птицы и возвращать мяч мне.
– Грач (грачата).
– Скворец (скворчата).
– Ласточка (ласточата).
– Дрозд (дроздята).
– Стриж (стрижата).
– Журавль (журавлята).
– Кукушка (кукушата).
Итог занятия. [Оценка работы детей] Воспитатель убирает все картинки и предлагает детям вспомнить, чем они сегодня занимались на занятии, в какие игры играли, что им понравилось делать.
перелетные птицы
перелетные птицы- Главная
- Ключевые слова
- перелетные птицы
-
Конспект НОД по ознакомлению с окружающим миром «Перелетные птицы» в старшей группе 2022
Автор: Корнеева Валентина Владимировна
Цель занятия: показать изменения жизни и поведения птиц с приходом весны.
перелетные птицы, старшая группа
-
Групповое коррекционно-развивающее занятие для обучающихся 3-го класса с задержкой психического развития по теме «Зимующие и перелетные птицы».
 2021
2021
Автор: Андреева Елена Юрьевна
Цель занятия: познакомить детей с перелетными птицами, развитие связной речи, зрительного внимания, мышления, артикуляционной, координации речи с движением.
зимующие птицы, ЗПР, перелетные птицы
-
Здравствуйте, птицы! 2021
Автор: Зарипова Рамиля Равиловна
Цель занятия: повысить знания детей о птицах; дать представление о перелетных птицах; выучить новые слова.

птицы, перелетные птицы
-
Конспект занятия в средней группе “Перелетные птицы” 2020
Авторы: Додонова Елена Васильевна, Моисеева Анна Михайловна
Занятие состоит из двух частей: ознакомительная информация о перелетных птицах и особенностях их образа жизни (грач, скворец, ласточка, аист, кукушка, соловей). Первые слайды знакомят детей с внешним видом и интересной информацией про птиц. Дают представление о звуках, которые издает каждая птица.
Задания способствуют развитию исследовательского поведения детей. Игры на закрепление и активизацию использования новых понятий в речи. Задания способствуют развитию связной речи (согласование числительного с существительным, использование предлогов в речи, умение аргументировать ответ, строить фразу при высказывании)
средняя группа, перелетные птицы
-
О чем поют весенние птицы 2020
Авторы: Бухер Галина Николаевна, Сухорукова Елена Павловна
Вместе с первым весенним теплом птицы летят на родину, на места своих прежних летних гнездовий.
 Они возвращаются друг за другом, начиная с марта и заканчивая началом мая.
Они возвращаются друг за другом, начиная с марта и заканчивая началом мая.
окружающий мир, литература, птицы, перелетные птицы
-
Конспект занятия “Перелётные птицы” в средней группе детского сада 2019
Автор: Ванюшкина Наталия Николаевна
Цели и задачи занятия: дать представление о перелетных птицах, познакомить с их внешним видом, образом жизни; закладывать основы экологического воспитания; активизировать наблюдательность.
весна, загадки,
Не нашли интересующий материал? Воспользуйтесь поиском по сайту.
Дальние мигрирующие виды птиц, путешествующие большими группами
Biol Lett. 2011 23 октября; 7(5): 692–694.
Опубликовано в сети 27 апреля 2011 г. doi: 10.1098/rsbl.2011.0243
Информация об авторе Примечания к статье Информация об авторских правах и лицензиях Отказ от ответственности
То, как люди мигрируют на большие расстояния, является вечной загадкой миграции животных. Было высказано предположение о сильном давлении отбора для путешествий группами у видов, мигрирующих на большие расстояния. Путешествие в группах может снизить энергетические потребности при длительной миграции, повысить точность навигации и способствовать групповому поиску пищи на миграционных стоянках. Тем не менее эта гипотеза не получила должного внимания. Я исследовал эволюционные переходы в дальности миграции у всех гнездящихся в Северной Америке видов птиц. Я задокументировал 72 эволюционных изменения расстояния миграции в пуле из 409разновидность. В противоположных кладах дальняя миграция, в отличие от миграции на короткие расстояния, была связана с большим размером путешествующей группы. Никаких других переходов не произошло наряду с другими признаками, такими как размер группы в негнездовой период или масса тела. Результаты показывают, что большие размеры групп были полезны в эволюции миграции на большие расстояния в большой кладе птиц.
Путешествие в группах может снизить энергетические потребности при длительной миграции, повысить точность навигации и способствовать групповому поиску пищи на миграционных стоянках. Тем не менее эта гипотеза не получила должного внимания. Я исследовал эволюционные переходы в дальности миграции у всех гнездящихся в Северной Америке видов птиц. Я задокументировал 72 эволюционных изменения расстояния миграции в пуле из 409разновидность. В противоположных кладах дальняя миграция, в отличие от миграции на короткие расстояния, была связана с большим размером путешествующей группы. Никаких других переходов не произошло наряду с другими признаками, такими как размер группы в негнездовой период или масса тела. Результаты показывают, что большие размеры групп были полезны в эволюции миграции на большие расстояния в большой кладе птиц.
Ключевые слова: птицы, формирование полета, численность группы, миграция, точность навигации
Миграция многих видов животных, от насекомых до птиц и крупных млекопитающих, часто связана с перемещениями на большие расстояния, в том числе захватывающими поездками на расстояние более 20 000 км у некоторых видов птиц [1].
Считается, что перемещение птиц стаями противодействует негативным последствиям миграции на большие расстояния с помощью трех различных механизмов. Во-первых, формирование стаи может улучшить аэродинамические характеристики, что дает значительную экономию энергии на больших расстояниях, особенно для более крупных видов [5]. Флокирование также может повысить способность обнаруживать неравномерно распределенные термики, что снижает транспортные расходы парящих видов [6].
Во-вторых, объединяя информацию о направлениях, стайки могут позволить птицам чаще двигаться в правильном направлении [7], а также могут помочь в обнаружении ограниченных или скрытых ориентиров во время полета [8]. Стайность также может дать неопытным людям возможность следовать за более знающими компаньонами к удаленной цели [9].,10].
Стайность также может дать неопытным людям возможность следовать за более знающими компаньонами к удаленной цели [9].,10].
В-третьих, передвижение стаями может способствовать формированию стаи во время миграционных остановок, поскольку птицы уже находятся в стаях, когда они останавливаются [11]. Было показано, что стаи во время кормления повышают эффективность кормодобывания и снижают риск хищничества [12] и могут быть особенно полезны в местах остановки, которые в лучшем случае посещаются редко [13].
Учитывая эти преимущества, у видов, мигрирующих на большие расстояния, должно существовать сильное давление отбора для перемещения в стаях [14,15]. Тем не менее мне известна только одна проверка гипотезы на одной кладе птиц [16]. Однако в этом исследовании не учитывался тот факт, что повторяющиеся случаи стада у мигрирующих на большие расстояния видов могут не быть независимыми эволюционными событиями, а вместо этого отражать наследование от общего предка. Здесь я исследовал эволюционные переходы в дальности миграции у всех гнездящихся в Северной Америке видов птиц.
Эволюционные переходы на расстояние миграции могут сопровождаться другими переходами, которые сами по себе могут объяснить какую-либо связь со стаями. Например, виды, мигрирующие на большие расстояния, могут быть более общительными в негнездовой период, чтобы повысить эффективность поиска пищи или снизить риск нападения хищников. Путешествие стаями будет просто отражать более общительный характер таких видов, который эволюционировал, чтобы приносить пользу не в первую очередь во время миграции, а в период, когда не размножается. Таким образом, я исследовал, собирают ли виды, мигрирующие на большие расстояния, в большие группы в негнездовой период. Точно так же я определил, что виды, мигрирующие на большие расстояния, как правило, меньше, что увеличило бы их относительный риск хищничества [17], а также способствовало бы большей общительности в период отсутствия размножения.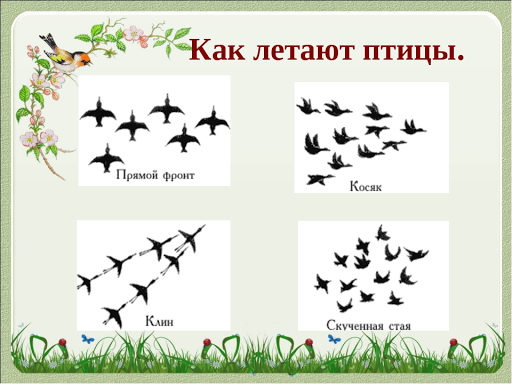 Я также рассматривал переходы к ночной миграции, которые могут препятствовать формированию стай [18].
Я также рассматривал переходы к ночной миграции, которые могут препятствовать формированию стай [18].
Я просмотрел литературу по птицам, чтобы определить склонность к миграции всех видов североамериканских птиц, гнездящихся на континенте. Миграция определялась как регулярные возвратные перемещения из года в год между местами размножения и зимовки. Я исключил виды с недостаточной информацией о миграции, инвазивные виды (нерегулярно перемещающиеся после размножения на разные расстояния), дисперсные виды (двигающиеся в любом направлении, включая высотных мигрантов) и неместные виды.
Я провел различие между видами, мигрирующими на короткие и дальние расстояния, в зависимости от того, проходил ли один этап миграционного пути не более 15° широты или более. Этого было достаточно в Северной Америке, поскольку большинство видов перемещаются по оси север-юг. Я отметил для каждого вида массу тела самца (МТ) (предпочтительно в негнездовой период), максимальный размер группы во время кормления в негнездовой период и максимальный размер группы во время полета во время миграции. Я подсчитывал максимальный размер группы во время поиска пищи, включая или исключая представителей других видов, поскольку многие виды добывали пищу в смешанных группах. Это различие редко применялось к размеру группы во время полета и не отмечалось. Максимальный размер группы сообщается чаще, чем другие статистические данные о группировке, и ранее был связан с экологическими особенностями [19].]. Я определил, происходила ли миграция преимущественно днем, а не днем и ночью.
Я подсчитывал максимальный размер группы во время поиска пищи, включая или исключая представителей других видов, поскольку многие виды добывали пищу в смешанных группах. Это различие редко применялось к размеру группы во время полета и не отмечалось. Максимальный размер группы сообщается чаще, чем другие статистические данные о группировке, и ранее был связан с экологическими особенностями [19].]. Я определил, происходила ли миграция преимущественно днем, а не днем и ночью.
Все количественные данные были логарифмически 10 преобразованы до статистического анализа. Филогенетический анализ этих количественных признаков основывался на независимых контрастах, рассчитанных с использованием модуля PDAP [20] в Mesquite [21], присваивая всем длинам ветвей значение 1. Филогения была основана на недавних работах, описывающих филогенетические отношения между птичьими семействами и внутри них (электронная дополнительная материал, приложение 1). Я полагался на реконструкцию состояния предков в пределах Мескита, чтобы установить наиболее экономное распределение категориальных признаков по филогенетическим линиям. Эта реконструкция позволила мне определить все эволюционные переходы в дальности миграции. Ни один из путей, соединяющих две линии с таким переходом, не перекрывал друг друга в целях обеспечения статистической независимости [22]. Я использовал ранговый критерий со знаком Уилкоксона для количественных переменных, чтобы установить, значительно ли отличалось от нуля распределение значений контраста для этих эволюционных переходов. Я использовал тест Макнемара [23], чтобы определить, были ли переходы между состояниями времени миграции более вероятными в том или ином направлении при смещении расстояния миграции.
Эта реконструкция позволила мне определить все эволюционные переходы в дальности миграции. Ни один из путей, соединяющих две линии с таким переходом, не перекрывал друг друга в целях обеспечения статистической независимости [22]. Я использовал ранговый критерий со знаком Уилкоксона для количественных переменных, чтобы установить, значительно ли отличалось от нуля распределение значений контраста для этих эволюционных переходов. Я использовал тест Макнемара [23], чтобы определить, были ли переходы между состояниями времени миграции более вероятными в том или ином направлении при смещении расстояния миграции.
Я обнаружил 72 эволюционных перехода в дальности миграции среди 409 видов североамериканских птиц, включенных в набор данных (электронный дополнительный материал, приложение 2). Эволюционные переходы охватили широкий круг семейств от уток до воробьев. Экономная реконструкция состояния показала, что расстояние миграции было коротким в основании дерева, и что эволюционные переходы в расстоянии миграции включали примерно одинаково часто переходы от короткой к дальней миграции или наоборот (электронный дополнительный материал, приложение 2).
В кладах с эволюционным переходом по расстоянию миграции максимальные размеры групп при полете были больше у видов, мигрирующих на большие расстояния ( S = −112,5, p < 0,0001; ) без сопутствующих изменений в BM ( S = 216,5, p = 0,17), максимальный размер группы сородичей во время поиска пищи ( S = -177,5, p = 0,17) или максимальный общий размер группы во время поиска пищи ( S = -128,5, p = 0,27). Наряду с эволюционными изменениями в расстоянии миграции время миграции менялось пять раз без какого-либо предпочтительного направления (9).0037 х 2 = 0,8, р = 0,37).
Открыть в отдельном окне
Блочные диаграммы филогенетически независимых контрастов по массе тела (BM), максимальному размеру группы в нерепродуктивный период (моноспецифический (MMGS) и гетероспецифический (MHGS)) и размеру путешествующей группы (MTGS) для мигрирующих видов североамериканских птиц. Отрицательные значения контраста указывают на то, что виды, мигрирующие на короткие расстояния, имеют более низкие значения, чем виды, мигрирующие на большие расстояния.
Отрицательные значения контраста указывают на то, что виды, мигрирующие на короткие расстояния, имеют более низкие значения, чем виды, мигрирующие на большие расстояния.
Независимые эволюционные переходы в дальности миграции были обычным явлением для большого пула североамериканских видов. Сходное открытие было сделано у двух родов европейских воробьиных птиц [24], что позволяет предположить, что миграция на большие расстояния у птиц несколько раз развивалась независимо [25]. Как и ожидалось, миграция на большие расстояния, в отличие от миграции на короткие расстояния в контрастных кладах, была связана с большим размером путешествующей группы.
Альтернативная гипотеза о том, что другие эволюционные переходы, происходившие одновременно с изменением расстояния миграции, объясняли увеличение размера передвижной стаи, кажется маловероятной. Изменение расстояния миграции не было связано с изменением размера негнездовой группы или BM, которые могли бы объяснить изменения в склонности к стаям во время путешествия. Стайность в негнездовой период также не была связана с расстоянием миграции в одной кладе птиц [26]. Точно так же произошло несколько изменений во времени миграции, что, как известно, влияет на стадо [18].
Стайность в негнездовой период также не была связана с расстоянием миграции в одной кладе птиц [26]. Точно так же произошло несколько изменений во времени миграции, что, как известно, влияет на стадо [18].
Недавние исследования показывают, что виды, ведущие оседлый образ жизни, а не мигрирующие, имеют больший относительный размер мозга, что будет способствовать инновациям в более суровых условиях, с которыми сталкиваются немигрирующие виды [27]. Можно утверждать, что виды, мигрирующие на большие расстояния, также выиграют от большей поведенческой гибкости из-за более серьезных проблем, с которыми сталкиваются виды, вынужденные добывать корм в самых разных местах обитания вдоль маршрута [4]. Поэтому в будущем было бы интересно связать относительный размер мозга с расстоянием миграции, а не только со склонностью к миграции вообще, и выяснить, специализируются ли одни люди на получении информации о направлении, в то время как другие используют информацию, предоставленную другими [15].
Смысл этого исследования заключается в том, что эволюционные изменения в размере стаи во время путешествия были связаны с изменением расстояния миграции. Неясно, позволили ли изначально большие размеры стаи увеличить продолжительность миграции или более длительная миграция способствовала эволюции более крупных стад. Картирование склонности к стаям и расстояний миграции на филогенетическом дереве можно использовать для оценки причинно-следственной связи, как это было сделано для других признаков, которые эволюционировали вместе с расстоянием миграции [28].
Эти находки подтверждают более раннее обнаружение у хищных птиц того, что передвижение стаями более характерно для видов, мигрирующих дальше [16]. Здесь я распространяю эти результаты на все гнездящиеся в Северной Америке виды птиц, используя эволюционную схему, в которой я определил независимые эволюционные переходы в дальности миграции. Этот вывод можно было бы проверить в других районах, например, в западной Палеарктике, где расстояния миграции и тенденции к стаям хорошо задокументированы [29].
Миграция также известна для других таксонов, таких как млекопитающие, рептилии, насекомые и рыбы. Миграция на большие расстояния может происходить в одиночку [30] или в больших группах [31], и задача будущих исследований будет состоять в том, чтобы определить, когда эволюция миграции в группах более вероятна. Очевидно, что путешествие в группах не является необходимым для дальней миграции, но результаты этого исследования показывают, что существует сильное давление отбора для путешествия большими группами, когда миграция происходит на большие расстояния в большой кладе видов птиц.
1. Ньютон И. 2008. Экология миграции птиц. Лондон, Великобритания: Academic Press [Google Scholar]
2. Piersma T. 1998. Гибкость фенотипа во время миграции: оптимизация размера органов в зависимости от рисков и преимуществ заправки топливом и полета? Дж. Авиан Биол. 29, 511–520 10.2307/3677170 (doi:10.2307/3677170) [CrossRef] [Google Scholar]
3. Alerstam T. 2006. Противоречивые данные о навигации животных на большие расстояния. Science 313, 791–794 10.1126/science.1129048 (doi:10.1126/science.1129048) [PubMed] [CrossRef] [Google Scholar]
Science 313, 791–794 10.1126/science.1129048 (doi:10.1126/science.1129048) [PubMed] [CrossRef] [Google Scholar]
4. Cimprich D. A., Woodrey M. S., Moore F. R. 2005. Воробьиные мигранты реагируют на изменение риска нападения хищников во время остановки. Аним. Поведение 69, 1173–1179 10.1016/j.anbehav.2004.07.021 (doi:10.1016/j.anbehav.2004.07.021) [CrossRef] [Google Scholar]
5. Weimerskirch H., Martin J., Clerquin Y., Александр П., Жираскова С. 2001. Энергосбережение при построении полета. Nature 413, 697–698 10.1038/35099670 (doi:10.1038/35099670) [PubMed] [CrossRef] [Google Scholar]
6. Kerlinger P., Gauthreaux S.A. 1985. Сезонные сроки, географическое распределение и полетное поведение ширококрылых ястребов во время весенней миграции в южном Техасе: радиолокационное и визуальное исследование. Auk 102, 735–743 [Google Scholar]
7. Саймонс А. М. 2004. Много ошибок: преимущество групповой навигации. Тенденции Экол. Evol 19, 453–455 10.1016/j.tree.2004.07. 001 (doi:10.1016/j.tree.2004.07.001) [PubMed] [CrossRef] [Google Scholar]
001 (doi:10.1016/j.tree.2004.07.001) [PubMed] [CrossRef] [Google Scholar]
8. Hamilton WJ 1962. Доказательства, касающиеся функции ночных криков перелетных птиц. Кондор 64, 390–401 10.2307/1365441 (doi:10.2307/1365441) [CrossRef] [Google Scholar]
9. Биро Д., Самптер Д., Мид Дж., Гилфорд Т. 2006. От компромисса к лидерству в голубеведении. Курс. биол. 16, 2123–2128 10.1016/j.cub.2006.08.087 (doi:10.1016/j.cub.2006.08.087) [PubMed] [CrossRef] [Google Scholar]
10. Кузин И. Д., Краузе Дж., Франкс Н. Р. , Левин С. А. 2005. Эффективное лидерство и принятие решений в группах животных на ходу. Nature 433, 513–516 10.1038/nature03236 (doi:10.1038/nature03236) [PubMed] [CrossRef] [Google Scholar]
11. Piersma T., Zwarts L., Bruggemann J.H. 1990. Поведенческие аспекты отлета куликов перед дальними перелетами: стаи, вокализации, траектории полета и суточный ритм. Ardea 78, 157–184 [Google Scholar]
12. Краузе Дж., Ракстон Г. Д. 2002. Жизнь в группах. Оксфорд, Великобритания: Издательство Оксфордского университета [Google Scholar]
13. Немет З., Мур Ф. Р. 2007. Незнакомые места остановки и ценность социальной информации во время миграции. Дж. Орнитол. 148, С369–S376 10.1007/s10336-007-0209-2 (doi:10.1007/s10336-007-0209-2) [CrossRef] [Google Scholar]
Немет З., Мур Ф. Р. 2007. Незнакомые места остановки и ценность социальной информации во время миграции. Дж. Орнитол. 148, С369–S376 10.1007/s10336-007-0209-2 (doi:10.1007/s10336-007-0209-2) [CrossRef] [Google Scholar]
14. Hamilton WJ. 1967. Социальные аспекты механизмов ориентации птиц. Ориентация животных и навигация (изд. Storm RM), стр. 57–71. Corvallis, OR: Oregon State University Press [Google Scholar]
15. Гуттал В., Кузин И. Д. 2010. Социальные взаимодействия, использование информации и эволюция. коллективной миграции. проц. Натл акад. науч. USA 107, 16 172–16 177 10.1073/pnas.1006874107 (doi:10.1073/pnas.1006874107) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Керлингер П. 1989. Стратегии полета мигрирующих ястребов. Чикаго, Иллинойс: University of Chicago Press [PubMed] [Google Scholar]
17. Коэн Дж. Э., Пимм С. Л., Йодзис П., Садана Дж. 1993. Размеры тела животных-хищников и животных-жертв в пищевых сетях. Дж. Аним. Экол. 62, 67–78 10.2307/5483 (doi:10.2307/5483) [CrossRef] [Google Scholar]
62, 67–78 10.2307/5483 (doi:10.2307/5483) [CrossRef] [Google Scholar]
18. Ларкин Р. П., Сафони Р. Э. 2008. Доказательства совместной ночной миграции широко рассредоточенных птиц. интегр. Комп. биол. 48, 40–4910.1093/icb/icn038 (doi:10.1093/icb/icn038) [PubMed] [CrossRef] [Google Scholar]
19. Beauchamp G. 2010. Расслабленный риск хищничества снижает, но не устраняет социальность у птиц. биол. лат. 6, 472–474 10.1098/rsbl.2009.1063 (doi:10.1098/rsbl.2009.1063) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
20. Midford P. E., Garland T., Maddison W. P. 2003. PDAP пакет Mesquite версии 1.
21. Maddison W.P., Maddison D.R. Mesquite: модульная система для эволюционного анализа. 2010. Версия 2.73. См. http://mesquiteproject.org.
22. Рид А.Ф., Ни С. 1995. Вывод из бинарных сравнительных данных. Дж. Теор. биол. 173, 99–108 10.1006/jtbi.1995.0047 (doi:10.1006/jtbi.1995.0047) [CrossRef] [Google Scholar]
Нью-Йорк, штат Нью-Йорк: McGraw-Hill [Google Scholar]
24. Хелбиг А. Дж. 2003. Эволюция миграции птиц. В птичьей миграции (редакторы Бертольд П., Гвиннер Э., Зонненшайн Э.), стр. 3–20. Берлин, Германия: Springer [Google Scholar]
Хелбиг А. Дж. 2003. Эволюция миграции птиц. В птичьей миграции (редакторы Бертольд П., Гвиннер Э., Зонненшайн Э.), стр. 3–20. Берлин, Германия: Springer [Google Scholar]
25. Алерстам Т., Хеденстром А., Акессон С. 2003. Дальняя миграция: эволюция и детерминанты. Oikos 103, 247–260 10.1034/j.1600-0706.2003.12559.x (doi:10.1034/j.1600-0706.2003.12559.x) [CrossRef] [Google Scholar]
26. Boyle W. A., Conway 20 C. J. Зачем мигрировать? Проверка гипотезы эволюционного предшественника. Являюсь. Нац. 169, 344–359 10.1086/511335 (doi:10.1086/511335) [PubMed] [CrossRef] [Google Scholar]
поведение птиц умеренной Палеарктики. проц. Р. Соц. Б 272, 1433–1441 10,1098/rspb.2005.3099 (doi:10.1098/rspb.2005.3099) [PMC free article] [PubMed] [CrossRef] [Google Scholar]
28. Правосудов В.В., Санфорд К., Хан Т.П. 2007. Об эволюции размера мозга в связи с миграционным поведением птиц. Аним. Поведение 73, 535–539 10.1016/j.anbehav.2006.10.005 (doi:10.1016/j.anbehav.2006.10.005) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Møller A. P. 2007. Старение в зависимости от широты и миграции у птиц. Дж. Эвол. биол. 20, 750–757 10.1111/j.1420-9101.2006.01236.x (doi:10.1111/j.1420-9101.2006.01236.x) [PubMed] [CrossRef] [Google Scholar]
Møller A. P. 2007. Старение в зависимости от широты и миграции у птиц. Дж. Эвол. биол. 20, 750–757 10.1111/j.1420-9101.2006.01236.x (doi:10.1111/j.1420-9101.2006.01236.x) [PubMed] [CrossRef] [Google Scholar]
KW, Thorrold S.R. 2009. Трансэкваториальные миграции гигантских акул в западной части Атлантического океана. Курс. биол. 19, 1019–1022 10.1016/j.cub.2009.04.019 (doi:10.1016/j.cub.2009.04.019) [PubMed] [CrossRef] [Google Scholar]
31. Mduma S.A.R., Sinclair A.R.E., Hilborn R. 1999. Еда регулирует гну Серенгети: 40-летний рекорд. Дж. Аним. Экол. 68, 1101–1122 10.1046/j.1365-2656.1999.00352.x (doi:10.1046/j.1365-2656.1999.00352.x) [CrossRef] [Google Scholar]
CSU AeroEco Lab Research Highlights
AeroEco Lab Research Highlights
Как изменилась фенология миграции за последние 20 с лишним лет?
Abstract: Городские зеленые насаждения (ПХГ) часто являются местом обитания разнообразных орнитологических сообществ. Текущие данные свидетельствуют о том, что большие ПХГ круглой формы с обильным пологом деревьев будут поддерживать большее количество видов птиц независимо от изоляции ПХГ. Здесь мы используем информацию о встречаемости птиц из научной базы данных eBird за 18-летний период (2002–2019 гг.).) для оценки годовых и сезонных моделей разнообразия видов птиц в хорошо обследованных зеленых насаждениях Нью-Йорка. Площадь была самым сильным коррелятом годового ( n = 102 UGS) и сезонного видового богатства ( n = 43 UGS), тогда как форма и изоляция не имели значимых связей. Увеличение площади на 50 % привело к увеличению годового видового богатства на 11,5 % и увеличению сезонного видового богатства на 8,2 %. Не было никаких доказательств того, что эти отношения содержали верхний предел. Покрытие древесного полога слабо коррелировало с годовым видовым богатством. Обилие ночных мигрирующих видов сильно коррелировало с площадью весной и осенью, особенно для неворобьиных видов. Видовое богатство ночных воробьиных воробьиных сильно коррелировало с древесным покровом весной, когда увеличение древесного покрова на 50% привело к увеличению видового богатства на 23,3%.
Текущие данные свидетельствуют о том, что большие ПХГ круглой формы с обильным пологом деревьев будут поддерживать большее количество видов птиц независимо от изоляции ПХГ. Здесь мы используем информацию о встречаемости птиц из научной базы данных eBird за 18-летний период (2002–2019 гг.).) для оценки годовых и сезонных моделей разнообразия видов птиц в хорошо обследованных зеленых насаждениях Нью-Йорка. Площадь была самым сильным коррелятом годового ( n = 102 UGS) и сезонного видового богатства ( n = 43 UGS), тогда как форма и изоляция не имели значимых связей. Увеличение площади на 50 % привело к увеличению годового видового богатства на 11,5 % и увеличению сезонного видового богатства на 8,2 %. Не было никаких доказательств того, что эти отношения содержали верхний предел. Покрытие древесного полога слабо коррелировало с годовым видовым богатством. Обилие ночных мигрирующих видов сильно коррелировало с площадью весной и осенью, особенно для неворобьиных видов. Видовое богатство ночных воробьиных воробьиных сильно коррелировало с древесным покровом весной, когда увеличение древесного покрова на 50% привело к увеличению видового богатства на 23,3%. Наши результаты подчеркивают большое значение территории, ценность древесного покрова для весенних мигрантов и ограниченное значение формы и изоляции. Усилия по увеличению площади и древесного покрова приведут к увеличению числа постоянных и мигрирующих видов птиц, что, вероятно, приведет к увеличению числа других чувствительных к площади лесных таксонов, встречающихся в городской среде.
Наши результаты подчеркивают большое значение территории, ценность древесного покрова для весенних мигрантов и ограниченное значение формы и изоляции. Усилия по увеличению площади и древесного покрова приведут к увеличению числа постоянных и мигрирующих видов птиц, что, вероятно, приведет к увеличению числа других чувствительных к площади лесных таксонов, встречающихся в городской среде.
Abstract: Наличие подробной информации, которая охватывает географический ареал вида, охватывает долгосрочный временной диапазон и дает информацию об отдельных людях (например, возраст и пол), является принципиальной проблемой в экологии. С этой целью Североамериканская лаборатория кольцевания птиц поддерживает уникальный и малоиспользуемый набор данных, который можно использовать для решения основных вопросов фенологических изменений у перелетных птиц. Мы использовали записи с 1966 по 2015 год, чтобы количественно определить, как изменились сроки миграции у дальних мигрантов, чернозобой голубой камышевки (9). 0037 Setophaga caerulescens ). Дополнительно изучались возрастные и половые различия в сроках миграционных перемещений. Мы заметили, что ранние весенние мигранты проходили через участки примерно на 1,1 дня раньше за десятилетие, и пик весенней миграции также приходился раньше за 50 лет этого исследования. Кроме того, фенологические изменения были более быстрыми с увеличением широты во время пиковых периодов весенней миграции. Осенью пик миграции оставался постоянным на протяжении всех 50 лет изучения, но в целом миграционный сезон показал затяжку. Весной самцы стабильно мигрировали раньше самок, а взрослые особи мигрировали раньше молодых особей. Осенью не было разницы во времени между самцами и самками, но молодые птицы мигрировали раньше, чем взрослые. Кроме того, весной миграция шла быстрее, чем осенью. Это исследование выявляет различные стратегии в отношении сроков миграции в зависимости от времени года, возрастных групп и пола, а также показывает, что корректировки в пути в зависимости от широты могут объяснять изменения в сроках миграции. Эта базовая информация о таком фундаментальном экологическом процессе имеет решающее значение для нашего понимания миграции, и мы должны использовать эти уникальные данные, чтобы оценить критические сдвиги в соответствующих масштабах миграции.
*внесли равный вклад в эту работу
Значение: Искусственный свет в ночное время является новым стимулом в истории эволюции ночных животных. Световое загрязнение может значительно изменить поведение этих организмов, от миграции до поиска пищи и голосового общения. Ночные мигрирующие птицы особенно восприимчивы к искусственному свету из-за адаптации и потребности в навигации и ориентировании в темноте. Однако влияние света на поведение в полете недостаточно точно определено, особенно в урбанизированной среде. Здесь мы сообщаем, что культовая городская световая инсталляция резко изменила поведение многих ночных мигрирующих птиц, но эти эффекты исчезли, когда свет был погашен. Мы рекомендуем выборочное удаление светового загрязнения в ночное время со значительной миграцией птиц, чтобы смягчить негативное воздействие на птиц, в частности столкновения с освещенными конструкциями.
Abstract: Существует несколько легкодоступных инструментов дистанционного зондирования для изучения ночных летающих животных (например, мигрирующих птиц), каждый из которых обладает уникальными ошибками измерений. Мы использовали три инструмента (радар наблюдения за погодой, тепловизионную инфракрасную камеру и акустический регистратор) для измерения временных и пространственных характеристик оценок ночного движения летающих животных весной и осенью 2011 и 2012 годов в Льюисе, штат Делавэр, США. Наша цель состояла в том, чтобы сравнить показатели различных технологий, чтобы лучше понять их предвзятость при обнаружении животных. Для радаров и тепловизоров наибольшая наблюдаемая интенсивность движения, как правило, приходилась на вечерние сумерки или вскоре после них, в то время как для акустического самописца пиковая активность птиц по крику полетов наблюдалась непосредственно перед утренними сумерками. Сравнивая интенсивность движения в ночное время для всех сезонов, мы обнаружили, что средние ночные корреляции между акустикой и двумя другими инструментами были слабо коррелированы (тепловизорная камера и акустика, r = 0,004 ± 0,04 SE, n = 100 ночей; радар и акустика, r = 0,14 ± 0,04 SE, n = 101 ночь), но сильно варьируется в зависимости от каждой ночи (диапазон = от -0,84 до 0,9). 2, диапазон = от -0,73 до 0,94). Средние ночные корреляции между интенсивностью движения, оцененной с помощью радара и тепловизионной инфракрасной камеры в течение ночи, имели более сильную положительную корреляцию (r = 0,39 ± 0,04 SE, n = 125 ночей), но также сильно различались для отдельных ночей (диапазон = -0,76). до 0,98). Путем сравнения с данными радара среди многочисленных интервалов высот мы определили, что высота летающего животного над землей положительно влияет на тепловизионное изображение и отрицательно на обнаружение полетных сигналов. Кроме того, тепловизионные обнаружения уменьшались при наличии облачного покрова и увеличивались при средней скорости полета животных, тогда как акустические обнаружения не показали никакой связи с наличием облачного покрова, но уменьшались при увеличении скорости полета. Мы обнаружили, что методы выборки имеют положительную корреляцию при сравнении средней ночной скорости трафика в разные ночи. Сила этих корреляций обычно возрастала в течение ночи, достигая максимума за 2–3 часа до утренних сумерек. Принимая во внимание сходимость измерений с помощью различных инструментов в это время, мы предлагаем исследователям рассмотреть возможность полетов по отбору проб в часы перед утренними сумерками, когда различия из-за систематической ошибки обнаружения среди инструментов отбора проб, по-видимому, сведены к минимуму.
Abstract: Мониторинг миграции птиц может принимать различные формы; однако мониторинг активной ночной миграции наземных птиц ограничен несколькими методами. Ночные крики птиц в настоящее время являются единственным методом описания состава мигрантов на видовом уровне. Однако по мере развития этого метода требуется больше информации, чтобы понять источники различий в обнаружении вызовов. Кроме того, в нескольких исследованиях изучается, как вероятности обнаружения различаются в различных атмосферных условиях. Мы используем записи ночных полетов от пленников, чтобы изучить зависимость обнаружения полетов от температуры и влажности воздуха.