06.05.2023|
|
0 Comment
Дифференциация к т: Картотека упражнений на дифференциацию звуков К
Дифференциация звуков К – Т. | Учебно-методический материал (старшая группа):
Дифференциация
звуков К-Т
Коля считает капли. Ребёнок пальчиком показывает на капли и произносит КАП-КАП-КАП-КАП
На вокзал прибыли поезда. Они сигналят. Ребёнок показывает на поезд и проговаривает ТУ-ТУ-ТУ.
Игра «По дорожке пройди, что видел расскажи».
Ребёнок пальчиком показывает и называет КАП, ТУ и т. д.
Игра «Доскажи-ка буковку»
На конце слов потерялись буквы, тебе нужно помочь Коле и Толе закончить слова. Взрослый говорит начало слова, а ребёнок договаривает последнюю букву слова.
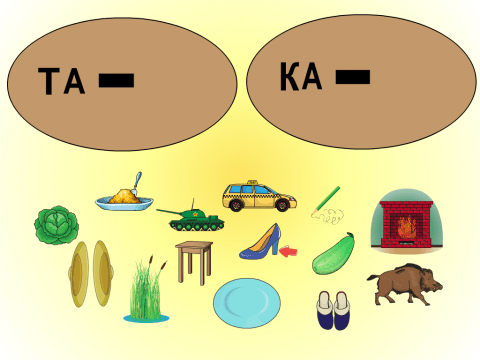
со_ живо_ бан_ лиф_ су_ щено_ лу_ кро_ клубо_ ко_ мешо_ жу_ гороше_ песо_ бегемо_ аис_ кус_
Дифференциация букв.
Толя очень любит ходить в гости к Коле. Толя прыгал по дороге с камушка на камушек. Прочитай буквы на камушках. (ребёнок пальчиком «прыгает» по камушкам и читает буквы)
Дифференциация в слогах.
Друзья любили зимой играть в снежки. Они налепили много снежков. Напиши на снежке букву «К», если услышишь слог со звуком «К». Напиши на снежке букву «Т», если услышишь слог со звуком «т». Посчитай, кто больше налепил снежков. Взрослый произносит слоги: КА, ТУ, КЫ, КО, ТА, ТЭ, АК, АТ и т.д, ребёнок записывает нужную букву в снежок. Потом читает «дорожку».
Повторение по одному, по два и по три слога со звуками К-Т.
ка – та – ка та-та – ка
та-ка, ка-та, ко-то, то-ко, ту-ку, ку-ту,
ат-ак, ат-ак, ут-ук, ук – ут
то-то-ко, ту-ту-ку, ко-то-ко, то-то-ко,
ак-ат-ак, ят-як-ят, от-ок-ок, ит-ик-ик, ик-ит-ик
ты-кы-кы, ка-та-ка, ку-ку-ту,
ака-ата, ото-око, ита-ика,
тла-кла, анк-ант, инк-инт, фта-фка и тд.
Предметы рассыпались. Расставь их по своим местам. Подбери слово, которое подойдет к схеме.
Дифференциация в словах.
Игра «Собери бусы»
Толя и Коля решили собрать бусы для мамы. У Коли были красные бусины, а у Толи – жёлтые.
У Коли были красные бусины, а у Толи – жёлтые.
Если услышишь слово со звуком «К», нанизывай красную бусину на ниточку. Если услышишь звук «Т», нанизывай жёлтую бусину.
Можно на столе выкладывать красные и жёлтые кружочки.
Можно на бумаге или доске рисовать и раскрашивать бусины.
Игра «Полёт на шаре»
Нужно пролететь вместе с шариком по дорожке и правильно назвать картинку со звуком Т или К. Можно добавить задание положить на картинку кружочек с нужной буквой.
Игра «Внимательные ушки».
Я буду произносить слова, а ты хлопни в ладоши, когда услышишь звук К, и топни ножками, когда услышишь звук Т.
Игра «Кто что делает».
Называй, кто это и что он делает.
Игра «Разложи картинки».
Коля и Толя любят собирать в свои альбомы картинки. Толя со звуком «Т», а Коля со звуком «К». Проведи стрелочки к альбомам.
Игра «Раскраска».
У Коли есть красный, коричневый, а у Толи жёлтый, фиолетовый.
Назови предметы и раскрась их этими карандашами.
Проговаривай предмет и его цвет: жёлтая лента, красная крыша.
Игра «Бродилка»
Проговаривание слов, в которых встречаются оба звука.
По дорожкам пройди и слова назови.
(кофта, конфета, комета, копыта, катушка, утка, ветка, космонавт, молоток, платок, моток, каток, цветок, танк, канат, охотник, капитан, трактор)
Дифференциация в словосочетаниях.
Игра «Поиграем посчитаем».
Посчитай диких уток, жёлтые цветки, маленькие улитки, вкусные конфеты.
Дифференциация в предложении.
Игра «Пазлы»
Взрослый разрезает картинку и просит ребёнка собрать и назвать все получившиеся картинки. Можно придумать с каждым их ни слов предложения.
Дифференциация в стихотворениях.
Коротышка под кроватку
Спрятал торт и шоколадку,
И картошку, и котлетку,
И капустку, и конфетку…
Ты постой-ка, коротышка!
Завтрак твой утащит мышка!
За столиком — Танюшка,
На столике — катушка.
Катушка тонких ниток
И ткани есть кусок…
Ну кто подскажет Тане —
Что сшить из этой ткани?
В тарелку косточку кладу —
Тотошку завтракать веду!
На окно забрался кот,
Умный кот, но кот не тот.
Дифференциация звуков [к-т-г-д]
Звуки [К], [Т], [Г], [Д] усваиваются детьми к 2-3
годам. Это звуки “раннего онтогенеза”. В
литературе достаточно внимания уделено
постановке и автоматизации заднеязычных звуков
(В.В.Коноваленко, С.В.Коноваленко, И.С.Лопухина),
тогда как переднеязычные звуки [Т], [Д]
рассмотрены недостаточно (И.С.Лопухина). Это
возникает из-за того, что наиболее частым
является замена звуков заднеязычных на звуки [Т],
[Д]. При встрече же с ребенком в прямо
противоположной ситуации (замена звуков [Т], [Д] на
звуки [К], [Г]) возникают трудности в поиске
лексического материала сначала на автоматизацию
указанных переднеязычных звуков, а затем на их
дифференциацию со звуками заднеязычными.
Поэтому главной задачей данной работы мы считаем подбор лексического материала по указанной проблеме.
Постановка и автоматизация звуков [Т], [Д]
На данном этапе работы из речевого материала исключаются звуки [Г], [К].
1. Формирование восприятия звуков [Т], [Д].
– “Хлопни, когда услышишь слово: (суп, таз, хвост, рот, стол, дом, удав)”. Одно из данных слов называется вперемежку с любыми другими словами.
– “Хлопни, когда услышишь слог : (та, ту, от, ды, до)”. Один из данных слогов называется вперемежку с любыми другими слогами, не содержащими звуков [Г], [Д].
– “Хлопни, когда услышишь звук [Т] ([Д])”. Звуки называются вперемежку с любыми звуками, кроме [Г], [К].
2. Изолированное произношение звука [Т] ([Д]).
Многократное изолированное произношение звука
с опорой на символы (Фомичева): “Как стучат
колеса поезда? – т-т-т”, “Как стучит по дереву
дятел? – д-д-д”.
3. Автоматизация звуков в слогах.
| Та – ту – то | да – ду – до |
| Ты – тэ – та | ды – дэ – да |
| От – ат – ит | ды – да – до |
| Ут – ат – от | ду – ды – до |
4. Автоматизация звуков в словах.
Слова со звуком [Т]: там, таз, Тома, Толя, туфли,
лото, это, четыре, лисята, волчата, медвежата,
бельчата, ежата, телята, поросята, цыплята,
индюшата, зайчата, жеребята, салат, халат, рот,
вот, самолет, пилот, тут, салют, машет, машут, спит,
спят, пьет, пьют, хлопает, хлопают, строит, строят,
нюхает, нюхают, смотрит, смотрят, бежит, плачет,
плачут, сидит, сидят, ест, едят, поет, поют,
танцует, танцуют, везет, везут, нет, винт, туман,
петух.
Слова со звуком [Д]: да, дай, Даша, дача, дом, дуб, дым, душ, дупло, духи, думай, Вода, иду, пирамида, пойду, уйду, беда, свобода, борода, помада, победа, команда, наряды, следы, пруды, сады, посуда.
5. Автоматизация звуков в словосочетаниях и предложениях.
Чаще всего на данном этапе уже можно использовать речевой материал, содержащий одновременно звуки и [Т] и [Д].
Там Толя. Тут Тома. Надену твои туфли. Я думаю, Даня спит. Зайчата оставили следы. Мама берет помаду. В саду спят поросята. Папа завинчивает винт. Тома хлопает в ладоши. Жеребята везут дрова. У деда борода. Бельчата сидят в дупле. В душе идет вода. Тома и Толя достали лото. Друзья едят дыню. Я иду по мосту. Вон там фонтан.
6. Стихотворные тексты со звуками [Т], [Д].
У енота тетя Тоня.
Тетя Тоня в тине тонет.
– Тетя Тоня, не тони!
– Эй, тяни ее, тяни!
Вот идут мои друзья,
Подавать обед пора.
Дуб стоит у дома,
Дом стоит у дуба.
Кот сидит у дуба.
Кот сидит у дома.
Дома мама, дома папа,
Только Томы нету рядом.
– Найдите нашу Тому!
– Танцует там у дома.
Дед Данила дает дыню –
Вот и Дане, вот и Тане.
Дятел на дубу сидит,
Дятел ветви долбит.
Мама моет пол,
Папа чинит стол,
Я посуду убрала,
У нас дома чистота.
Даша и Наташа идут в лес. В лесу они видят дуб. На ветке дуба сидят бельчата. Даша и Наташа машут бельчатам: “Идите домой, в дупло!”
Толя идет в душ. Там льется вода. Толя стоит под душем. Потом он намыливается. Хорошо быть чистым и умытым!
Дифференциация звуков [К-Т]
А. Дифференциация звуков [К-Т] на слух.
– Игра “Хлопни-топни”. Инструкция: “Если
услышишь слог ТА – хлопни, если услышишь слог КА
– топни”.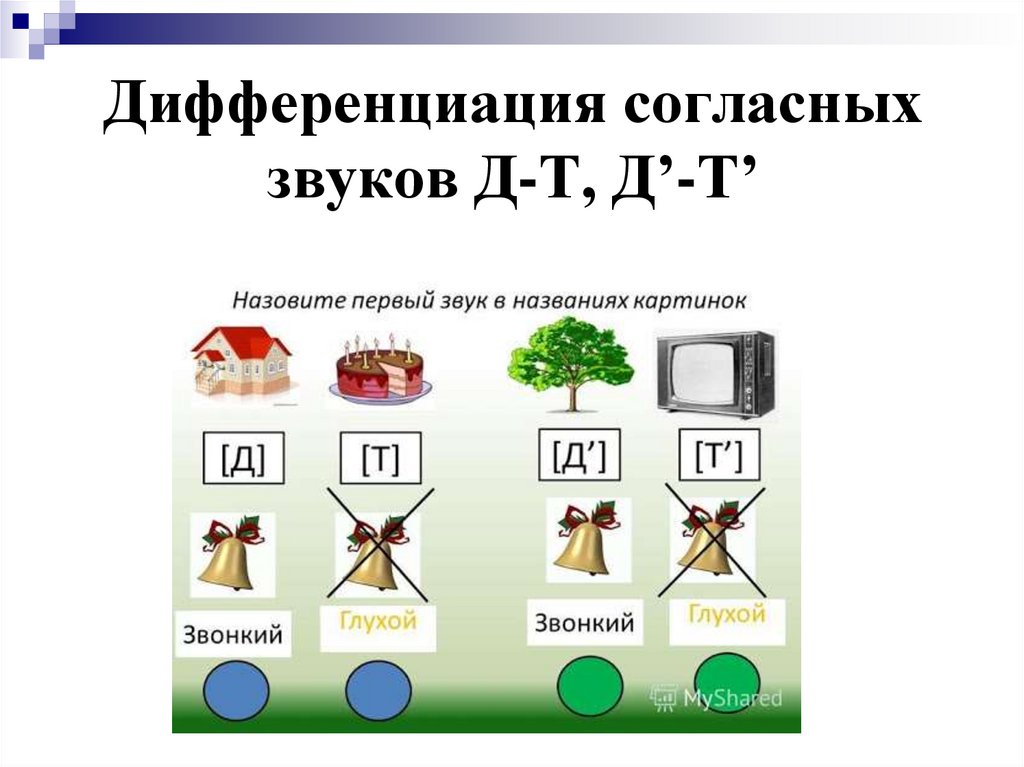 Позднее, игра проводится на материале
звуков. На этом этапе возможны любые
традиционные игры на дифференциацию звуков речи.
Позднее, игра проводится на материале
звуков. На этом этапе возможны любые
традиционные игры на дифференциацию звуков речи.
Б. Дифференциация произношения звуков [К-Т].
1. Попеременное называние изолированных звуков. Игра “Дождик-поезд”. Инструкция: “Когда я говорю дождик, ты произносишь звук [К], когда я говорю поезд, ты произносишь звук [Т]”.
2. Повторение по одному, по два и по три слога со звуками [К-Т].
ка – та – ка та – та – ка
ты – кы – ты ко – ко – то
ку – ту – ту ты – кы – кы
3. Повторение слов и называние картинок.
Словарь: кабинет, канат, капитан, капот, катать, катание, кафтан, ватка, ветка, детка, девятка, дудка, утка, кот, ток, комета, комната, комод, конфета, копыто, кофта, подкова, молоток, моток, охотник, тук, стук, путник, спутник, каток, конфета, тыква, свисток, селедка, лодка, платок.
4. Дифференциация звуков в словосочетаниях и
предложениях. Задания:
Задания:
а) “Посчитай до пяти”. Выбирается несколько слов из вышеуказанного словаря,
б) “Спроси и ответь”. Про любой неодушевленный предмет ребенок спрашивает и отвечает: “Это что? – Это канат”. Про любое животное ребенок спрашивает и отвечает: “Это кто? – Это собака” или “Кто тут? – Тут овца”.
Дифференциация звуков [Г-Д]
Задания, как и в предыдущем разделе.
Словарь: гадать, гадание, угадать, дуга, радуга, погода, ягода, когда, гнедой, города.
Дифференциация звуков [К-Т-Г-Д]
1. Словарь: отгадка, агат, Агата, тайга, гоготать, гонит, коготок, готов, фагот, команда, подруга, градусник, удочка, дождик, селедочка, гусята, лодочка.
2. Предложения:
Катя собирает букет. В комнате летает муха. В
доме два этажа. Коля ест торт. На столе стоит
кактус. Моряк смотрит в бинокль. Я иду в аптеку за
ватой. Таня надевает кофту. В комнате стоит комод.
Мама надевает платок. На тарелке лежат конфеты. Дед тянет репку. Высоко гнездо аиста. На
остановке стоит автобус. На окне сидит кот. Паук
плетет паутину. У Кати красивый бант. Дедушка
видит бегемота.
Дед тянет репку. Высоко гнездо аиста. На
остановке стоит автобус. На окне сидит кот. Паук
плетет паутину. У Кати красивый бант. Дедушка
видит бегемота.
3. Тексты:
Опускается туман
На мосты и на кусты,
Опускается туман
На дома и на сады,
Тут не видно далеко,
Заблудиться так легко!Соловей выводит трель.
Слесарь в сеть включает дрель.
Лом ломает толстый лед,
Лом ломать не устает.Вот так домик!
Вот так дом!
В доме том одно окно,
В том окне идет кино.– Тук! Тук! Тук!
– Это кто тут?
– Это я, коток,
Тонкий коготок.Всё готово для хоккея:
И коньки и капитаны,
На катке стоят команды,
Ждут свистка, как кот сметаны!-Я иду куда?
– Туда.
– Я пойду от кого?
– От того.
– Будет там кто?
– Не будет там никого.
– Не пойду туда,
Буду дома я тогда!Вот на стуле кот сидит,
Вот за бантом он следит.
Когда бантик двинется,
Он на бантик кинется!
Наступила зима. На дворе холодно, стужа. Деревья, крыши домов, дороги – все покрыто снегом. Реки покрыты льдом. Рады дети. Они бегут с коньками на каток.
Большинство приведенных текстов являются авторскими. Для самостоятельного подбора лексического материала по указанной проблеме можно воспользоваться литературой из списка.
Литература
- Коноваленко В.В., Клноваленко С.В. Дидактический материал по коррекции произношения звуков К, К’, Г, Г’, Х, Х’. – М.: “ГНОМ-ПРЕСС”, 1999.
- Лопухина И.С. Логопедия, упражнения для развития
речи: Пособие для логопедов и родителей. – СПб.:
Дельта, 1999.

- Смирнова М.В. 2500 скороговорок. Для детей 4-6 лет. - СПб.: Издательский дом “Нева”, 2005.
Дифференциация Т-клеток – Creative Diagnostics
Т-клетки (Т-лимфоциты) получили свое название от органов, в которых они развиваются, в тимусе. Они возникают в костном мозге, но для созревания мигрируют в вилочковую железу. Разнообразные реакции Т-клеток в совокупности называются клеточно-опосредованными иммунными реакциями. Это сделано для того, чтобы отличить их от ответов антител, которые, конечно же, также зависят от клеток (В-клеток). Т-клетки не могут распознавать антиген отдельно, так как рецепторы Т-клеток (TCR) могут распознавать только антиген, связанный с белками клеточной мембраны (молекулами MHC). TCR имеют разную структуру, поэтому они связываются с разными молекулярными структурами и имеют разные генетические коды. Подобно ответам антител, ответы Т-клеток исключительно антигенспецифичны и, по крайней мере, так же важны, как и антитела, в защите позвоночных от инфекций. Действительно, для инициации большинства адаптивных иммунных ответов, включая реакции антител, требуются Т-хелперы. Самое главное, в отличие от В-клеток, Т-клетки могут помочь устранить патогены, находящиеся внутри клеток-хозяев.
Действительно, для инициации большинства адаптивных иммунных ответов, включая реакции антител, требуются Т-хелперы. Самое главное, в отличие от В-клеток, Т-клетки могут помочь устранить патогены, находящиеся внутри клеток-хозяев.
Типы Т-клеток
В зависимости от функции и поверхностного маркера Т-клетки можно разделить на четыре основных класса.
- Цитотоксические Т-клетки непосредственно убивают инфицированные клетки, заставляя их подвергаться апоптозу, эти клетки подобны «киллерам» или цитотоксинам, потому что они убивают представляющие интерес клетки, продуцирующие определенный антиген. Основным поверхностным маркером цитотоксических Т-клеток является CD8, также известный как Т-клетки-киллеры.
- Т-хелперы играют промежуточную роль в иммунном ответе. Они пролиферируют, чтобы активировать В-клетки, чтобы вызвать реакцию антител, и макрофаги, чтобы уничтожить микроорганизмы, которые либо вторглись в макрофаги, либо были проглочены ими.
 Вспомогательные Т-клетки также помогают активировать цитотоксические Т-клетки для уничтожения инфицированных клеток-мишеней. Сами Т-хелперы, однако, могут функционировать только при активации, чтобы стать эффекторными клетками. Основным поверхностным маркером Т-хелперов является CD4.
Вспомогательные Т-клетки также помогают активировать цитотоксические Т-клетки для уничтожения инфицированных клеток-мишеней. Сами Т-хелперы, однако, могут функционировать только при активации, чтобы стать эффекторными клетками. Основным поверхностным маркером Т-хелперов является CD4. - Т-клетки памяти состоят как из CD4, так и из CD8 Т-клеток, которые могут быстро приобретать эффекторные функции для уничтожения инфицированных клеток и/или секретировать воспалительные цитокины, подавляющие репликацию патогена. Вместе с В-клетками памяти лимфоциты, которые сохраняют специфические антигенные сообщения после антигенной стимуляции, имеют продолжительность жизни до нескольких десятилетий. Когда они снова получают те же самые антигенные стимулы, они могут пролиферировать как функциональные Т-клетки против антигена или плазматических клеток, которые продуцируют антитела.
- Регуляторные/супрессорные Т-клетки часто играют важную роль в поддержании собственной толерантности и предотвращении чрезмерного повреждения иммунного ответа организма.
 Существует множество классов регуляторных/супрессорных Т-клеток, включая Т-клетки CD25 и CD4. Они могут ингибировать Т-клетки и В-клетки, чтобы регулировать и контролировать иммунный ответ и поддерживать самостабильность иммунной системы.
Существует множество классов регуляторных/супрессорных Т-клеток, включая Т-клетки CD25 и CD4. Они могут ингибировать Т-клетки и В-клетки, чтобы регулировать и контролировать иммунный ответ и поддерживать самостабильность иммунной системы.
Обзор дифференцировки Т-клеток
В тимусе развивающиеся Т-клетки, известные как тимоциты, пролиферируют и дифференцируются в соответствии с путями развития, которые генерируют функционально отличные субпопуляции зрелых Т-клеток. Помимо того, что он является основным источником всех Т-клеток, это место, где Т-клетки разнообразны, а затем формируются в эффективный первичный репертуар Т-клеток с помощью экстраординарной пары процессов отбора.
Дифференцировка клеток необходима для создания множества подмножеств. Дифференцировка наивных Т-клеток в эффекторные клетки необходима для оптимальной защиты от различных классов микробных патогенов и для развития иммунной памяти. Дифференцирующиеся клетки претерпевают запрограммированные изменения паттернов экспрессии генов, которые регулируются структурными изменениями хроматина.
Дифференцировка также управляется инструктивными и разрешающими сигналами из окружающей среды, особенно от антигенпрезентирующих клеток (АРС). Эти клетки определяют класс проглоченного микроба и генерируют сигналы, которые направляют наивные Т-клетки на дифференцировку в подмножество, которое мобилизует соответствующие механизмы иммунной защиты. Широко распространено мнение, что цитокины являются основными факторами дифференцировки. Однако модель, управляемую исключительно цитокинами, трудно согласовать с доказательствами того, что презентация антигена и доставка сигналов дифференцировки происходят с помощью одного и того же APC. Следовательно, решающую роль могут играть факторы ближнего действия, такие как молекулы клеточной поверхности.
Дифференциация Т-клеток CD4 и CD8
В последнее десятилетие было обнаружено постоянно растущее число субпопуляций Т-хелперных клеток CD4 с уникальными транскрипционными программами, управляемыми определяющими линию транскрипционными факторами. Подмножества Т-хелперных клеток CD4, известные как Т-хелперы 1 (Th2), Th3, Th9, Th27 и Th32, продуцируют определенные коктейли цитокинов для координации иммунитета к различным типам микроорганизмов. Фолликулярные Т-хелперы (Tfh) специализируются на помощи В-клеткам, в то время как индуцированные регуляторные Т-клетки (iTreg) подавляют пагубные иммунные реакции. Наконец, требуется этап дифференцировки для создания Т-клеток, которые способствуют немедленному отторжению микробной инфекции, а также других клеток, которые развиваются в клетки памяти.
Подмножества Т-хелперных клеток CD4, известные как Т-хелперы 1 (Th2), Th3, Th9, Th27 и Th32, продуцируют определенные коктейли цитокинов для координации иммунитета к различным типам микроорганизмов. Фолликулярные Т-хелперы (Tfh) специализируются на помощи В-клеткам, в то время как индуцированные регуляторные Т-клетки (iTreg) подавляют пагубные иммунные реакции. Наконец, требуется этап дифференцировки для создания Т-клеток, которые способствуют немедленному отторжению микробной инфекции, а также других клеток, которые развиваются в клетки памяти.
Т-хелперные клетки типа 1 (Th2) обычно транскрибируют фактор транскрипции T-bet и продуцируют гамма-интерферон (IFNg), интерлейкин-2 (IL-2) и гены лимфотоксина, тогда как клетки Th3 экспрессируют IL-4, IL -5, IL-6, IL-10 и IL-13. Как Th2, так и Th3 клетки, по-видимому, происходят от общих наивных клеток-предшественников, путь дифференцировки которых определяется цитокиновыми и костимулирующими сигналами во время первичной антигенной стимуляции. В частности, дифференцировка Th2 управляется IL-12 и требует IL-12-чувствительного транскрипционного фактора STAT4, в то время как Th3-дифференциация вызывается IL-4 и требует IL-4-чувствительного фактора транскрипции STAT6. In vivo прогрессивная поляризация цитокинового ответа возникает в ответ на хроническую антигенную стимуляцию в результате самоамплификации и негативной перекрестной регуляции, присущих дифференцировке Т-клеток. Клетки Th2 направляют иммунитет против внутриклеточных бактерий и вирусов и участвуют во множестве аутоиммунных состояний. Тем не менее, клетки Th3 развиваются у мышей, инфицированных гельминтами, даже если передача сигналов рецептора IL4 генетически отключена, что демонстрирует существование других путей, которые индуцируют эту программу дифференцировки in vivo . Ответы клеток Th3 также были снижены с помощью GSI в модели или при астме.
В частности, дифференцировка Th2 управляется IL-12 и требует IL-12-чувствительного транскрипционного фактора STAT4, в то время как Th3-дифференциация вызывается IL-4 и требует IL-4-чувствительного фактора транскрипции STAT6. In vivo прогрессивная поляризация цитокинового ответа возникает в ответ на хроническую антигенную стимуляцию в результате самоамплификации и негативной перекрестной регуляции, присущих дифференцировке Т-клеток. Клетки Th2 направляют иммунитет против внутриклеточных бактерий и вирусов и участвуют во множестве аутоиммунных состояний. Тем не менее, клетки Th3 развиваются у мышей, инфицированных гельминтами, даже если передача сигналов рецептора IL4 генетически отключена, что демонстрирует существование других путей, которые индуцируют эту программу дифференцировки in vivo . Ответы клеток Th3 также были снижены с помощью GSI в модели или при астме.
Пролиферация Т-клеток CD8 зависит от повторяющихся столкновений с антигеном. Каждая клетка, стимулированная антигеном, делится и постепенно дифференцируется в эффекторные цитотоксические Т-лимфоциты (ЦТЛ), а затем в CD8 Т-клетки памяти с каждым последующим клеточным делением. Первоначальный антигенный стимул запускает эту программу развития, так что Т-клетки CD8 начинают пролиферировать и дифференцироваться. Дальнейшая антигенная стимуляция дочерних клеток может увеличить число делений активированных CD8 T-клеток, но это не обязательно для завершения этой программы развития.
Первоначальный антигенный стимул запускает эту программу развития, так что Т-клетки CD8 начинают пролиферировать и дифференцироваться. Дальнейшая антигенная стимуляция дочерних клеток может увеличить число делений активированных CD8 T-клеток, но это не обязательно для завершения этой программы развития.
Запрограммированное развитие Т-клеток CD8 имеет несколько преимуществ. Во-первых, это устраняет необходимость длительного удержания ЦТЛ в лимфоидных органах, что позволяет им мигрировать к периферическим участкам инфекции и/или воспаления для удаления инфицированных клеток. Во-вторых, это также может значительно повлиять на количество генерируемых Т-клеток памяти CD8, поскольку размер пула Т-клеток памяти прямо коррелирует с размером популяции эффекторных клеток1-3. В некоторых моделях острой вирусной и бактериальной инфекции количество эффекторных CD8 Т-клеток достигает пика через 2–3 дня после элиминации инфекционного патогена. Если бы каждое деление Т-клеток CD8 строго регулировалось контактом с антигеном, число эффекторных ЦТЛ достигало бы пика раньше и достигало бы более низкого максимума, и, следовательно, генерировалось бы меньше Т-клеток памяти CD8.
Развитие ответов эффекторных Т-клеток тесно связано с клональной экспансией. Исследования показали, что связь между приверженностью к клональной экспансии и дифференцировкой эффекторных клеток очень тесная; та же продолжительность антигенной стимуляции (2–24 часа), которая стимулировала пролиферацию наивных CD8 T-клеток, была достаточной для их фиксации, чтобы дифференцироваться в эффекторные клетки, фактор некроза опухоли (TNF) и IL-2 и уничтожать инфицированные клетки.
Заболевания и лечение дифференцировки Т-клеток
Т-хелперные (Th)-лимфоциты проходят две пространственно и во времени различные фазы дифференцировки. После первой фазы развития, которая происходит в тимусе, вторая фаза, запускаемая начальным столкновением с антигеном на периферии, приводит к развитию субпопуляций эффекторных Т-хелперов, демонстрирующих взаимоисключающие паттерны экспрессии генов цитокинов. Клинически Th2-паттерны продукции цитокинов связаны с воспалением и аутоиммунными заболеваниями, тогда как Th3-паттерны характерны для аллергических реакций и астмы. Подобно системной красной волчанке (СКВ), это аутоиммунное заболевание неизвестной этиологии, поражающее более миллиона человек ежегодно. СКВ характеризуется гиперактивностью В- и Т-клеток и дефектами клиренса апоптотических клеток и иммунных комплексов. Понимание вовлеченного сложного процесса и взаимодействия между различными цитокинами, хемокинами, сигнальными молекулами и рецепторами распознавания образов (PRR) в иммунных путях предоставит ценную информацию о разработке новых терапевтических мишеней для лечения СКВ.
Подобно системной красной волчанке (СКВ), это аутоиммунное заболевание неизвестной этиологии, поражающее более миллиона человек ежегодно. СКВ характеризуется гиперактивностью В- и Т-клеток и дефектами клиренса апоптотических клеток и иммунных комплексов. Понимание вовлеченного сложного процесса и взаимодействия между различными цитокинами, хемокинами, сигнальными молекулами и рецепторами распознавания образов (PRR) в иммунных путях предоставит ценную информацию о разработке новых терапевтических мишеней для лечения СКВ.
Каталожные номера:
- Альбертс, Б.; и др. . Молекулярная биология клетки. 2002.
- Каеч, С.М.; и др. . Дифференцировка эффекторных и Т-клеток памяти: последствия для разработки вакцин. Природа . 2002, 2(4): 251-62.
Вернуться к ресурсам
Редакционная статья: Дифференцировка и функция Т-клеток при воспалении тканей
Т-клетки, составляющие одно из звеньев адаптивного иммунного ответа, обеспечивают клеточно-опосредованный иммунитет против патогенных микроорганизмов. Тимус является местом созревания Т-клеток, которые, как было показано, участвуют в клеточно-опосредованном иммунитете и гуморальном иммунном ответе у 19 животных.61–1962. Потребовалось еще несколько десятилетий, чтобы идентифицировать гетеродимерный рецептор Т-клеток, который имеет решающее значение для активации, дифференцировки и функций Т-клеток (1). В течение следующих 25–30 лет несколько новаторских исследований способствовали общему влиянию Т-клеток на модуляцию иммунных реакций в норме и при заболеваниях. Дифференцировка Т-клеток является одним из ключевых событий, которые абсолютно необходимы не только для элиминации внутри- и внеклеточных патогенов, но и при нарушении регуляции могут также привести к возникновению воспаления с обострением патогенеза заболевания при аутоиммунных заболеваниях. Эта тема исследования была разработана, чтобы понять сложность и молекулярные пути, которые приводят к дифференцировке Th-клеток, вызывающей патогенез заболевания. В рамках этой темы исследования была опубликована серия статей, в которых содержится содержательная информация об этой новой области.
Тимус является местом созревания Т-клеток, которые, как было показано, участвуют в клеточно-опосредованном иммунитете и гуморальном иммунном ответе у 19 животных.61–1962. Потребовалось еще несколько десятилетий, чтобы идентифицировать гетеродимерный рецептор Т-клеток, который имеет решающее значение для активации, дифференцировки и функций Т-клеток (1). В течение следующих 25–30 лет несколько новаторских исследований способствовали общему влиянию Т-клеток на модуляцию иммунных реакций в норме и при заболеваниях. Дифференцировка Т-клеток является одним из ключевых событий, которые абсолютно необходимы не только для элиминации внутри- и внеклеточных патогенов, но и при нарушении регуляции могут также привести к возникновению воспаления с обострением патогенеза заболевания при аутоиммунных заболеваниях. Эта тема исследования была разработана, чтобы понять сложность и молекулярные пути, которые приводят к дифференцировке Th-клеток, вызывающей патогенез заболевания. В рамках этой темы исследования была опубликована серия статей, в которых содержится содержательная информация об этой новой области.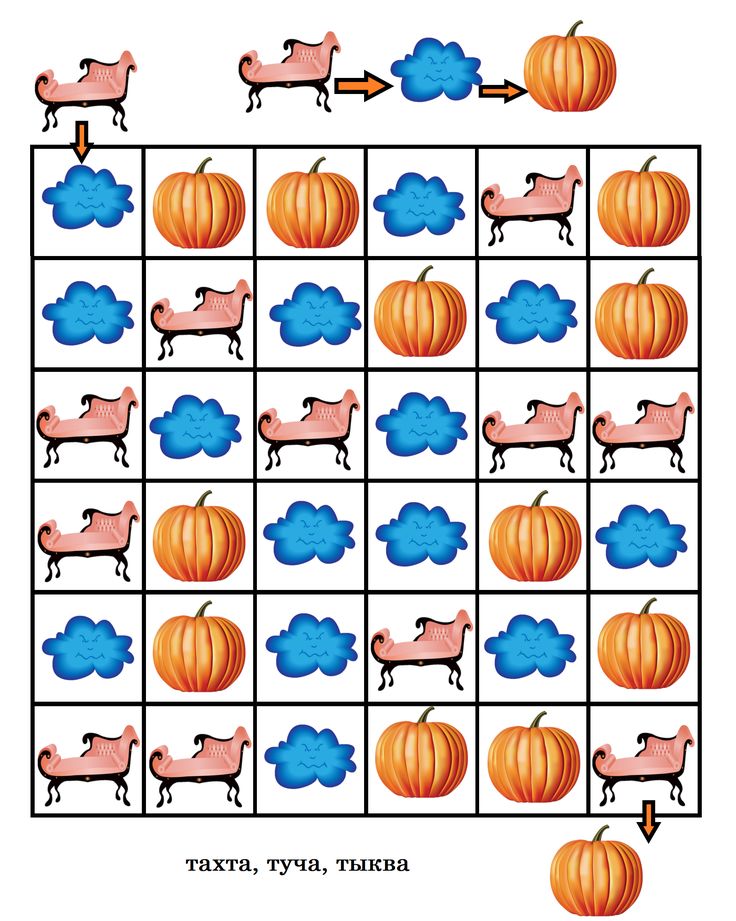 Вкратце, этот специальный выпуск состоит из 8 оригинальных исследовательских статей, 5 полноформатных обзоров, 3 мини-обзоров и 1 перспективы для обсуждения влияния активации и дифференцировки Т-клеток на воспаление тканей. Оригинальные исследовательские статьи включали роль CD4+ Т-клеток в патофизиологии неинфекционного увеита и болезни Грейвса. Многогранная роль различных субпопуляций CD4+ Т-клеток была подробно рассмотрена в гомеостазе тканей, воспалительных заболеваниях кишечника, остеопорозе и нейровоспалении. Эти статьи решительно поддерживают и дают новое представление о том, что использование знаний о дифференцировке Th-клеток может открыть новые терапевтические стратегии для контроля воспалительных заболеваний.
Вкратце, этот специальный выпуск состоит из 8 оригинальных исследовательских статей, 5 полноформатных обзоров, 3 мини-обзоров и 1 перспективы для обсуждения влияния активации и дифференцировки Т-клеток на воспаление тканей. Оригинальные исследовательские статьи включали роль CD4+ Т-клеток в патофизиологии неинфекционного увеита и болезни Грейвса. Многогранная роль различных субпопуляций CD4+ Т-клеток была подробно рассмотрена в гомеостазе тканей, воспалительных заболеваниях кишечника, остеопорозе и нейровоспалении. Эти статьи решительно поддерживают и дают новое представление о том, что использование знаний о дифференцировке Th-клеток может открыть новые терапевтические стратегии для контроля воспалительных заболеваний.
В то время как CD4+ T-клетки высвобождают цитокины, CD8+ T-клетки являются цитотоксическими. Недавнее исследование подкрепило мнение о том, что CD8+ Т-клетки могут быть важным фактором долголетия (2). Адоптивное лечение Т-клетками показало огромный потенциал для обучения иммунной системы борьбе со смертельными заболеваниями, такими как рак. Опухолеспецифические CD8+ Т-клетки вводятся пациентам, которые нацеливаются на раковые клетки и атакуют их. Существуют клинические испытания, которые показали успешный результат лечения метастатической меланомы с использованием адоптивной Т-клеточной терапии. Т-клетки пациентов были генетически модифицированы с помощью синтетических рецепторов, генерирующих химерные антигенные рецепторы Т-клеток (CAR T) для специфического нацеливания на поверхностный антиген раковых клеток. Для терапии CAR Т-клетками доступно несколько мишеней, включая иммуномодулирующие антигены (PD-L1), сверхэкспрессированные антигены (EGFR, HER2), аберрантно гликозилированные белки (MUC1). Планируется, что гены суицида будут включены в CAR Т-клетки, чтобы действовать как предохранительный выключатель.
Опухолеспецифические CD8+ Т-клетки вводятся пациентам, которые нацеливаются на раковые клетки и атакуют их. Существуют клинические испытания, которые показали успешный результат лечения метастатической меланомы с использованием адоптивной Т-клеточной терапии. Т-клетки пациентов были генетически модифицированы с помощью синтетических рецепторов, генерирующих химерные антигенные рецепторы Т-клеток (CAR T) для специфического нацеливания на поверхностный антиген раковых клеток. Для терапии CAR Т-клетками доступно несколько мишеней, включая иммуномодулирующие антигены (PD-L1), сверхэкспрессированные антигены (EGFR, HER2), аберрантно гликозилированные белки (MUC1). Планируется, что гены суицида будут включены в CAR Т-клетки, чтобы действовать как предохранительный выключатель.
Дифференцированные CD4+ Т-клетки играют решающую роль в обеспечении полезных иммунных ответов против патогенных микроорганизмов. И наоборот, CD4+ Т-клетки играют различные роли в патологии аутоиммунного воспаления. Эффекторные CD4+ Т-клетки, которые Mosmann et al. первоначально классифицировали как клетки Th2 и Th3. (3) расширились за последние 3 десятилетия с появлением клеток Th27, Th9, Tfh и Th32. Важно отметить, что CD4+ Т-клетки не только инициируют специфические иммунные ответы; также были идентифицированы субпопуляции CD4+ Т-клеток, которые способны ингибировать инициацию иммунных реакций и даже подавлять установленные иммунные ответы. Эти CD4+ Т-клетки называются регуляторными Т-клетками (Tregs) и из-за их роли в иммунопатогенезе аутоиммунных заболеваний и их потенциального использования в терапевтических целях стали предметом интенсивных исследований. Treg, секретирующие IL-10, были обозначены как клетки Tr1, которые не экспрессируют Foxp3. Наивные CD4+ Т-клетки могут найти свою нишу в воспаленных тканях при некоторых аутоиммунных заболеваниях, которые в противном случае были бы ограничены кровообращением и вторичными лимфоидными органами. Однако аллергическое воспаление в результате Th3-опосредованного ответа на аллергены окружающей среды и Th2-опосредованного иммунитета ответственно за возникновение множественных органоспецифических экспериментальных аутоиммунных заболеваний у животных.
Эффекторные CD4+ Т-клетки, которые Mosmann et al. первоначально классифицировали как клетки Th2 и Th3. (3) расширились за последние 3 десятилетия с появлением клеток Th27, Th9, Tfh и Th32. Важно отметить, что CD4+ Т-клетки не только инициируют специфические иммунные ответы; также были идентифицированы субпопуляции CD4+ Т-клеток, которые способны ингибировать инициацию иммунных реакций и даже подавлять установленные иммунные ответы. Эти CD4+ Т-клетки называются регуляторными Т-клетками (Tregs) и из-за их роли в иммунопатогенезе аутоиммунных заболеваний и их потенциального использования в терапевтических целях стали предметом интенсивных исследований. Treg, секретирующие IL-10, были обозначены как клетки Tr1, которые не экспрессируют Foxp3. Наивные CD4+ Т-клетки могут найти свою нишу в воспаленных тканях при некоторых аутоиммунных заболеваниях, которые в противном случае были бы ограничены кровообращением и вторичными лимфоидными органами. Однако аллергическое воспаление в результате Th3-опосредованного ответа на аллергены окружающей среды и Th2-опосредованного иммунитета ответственно за возникновение множественных органоспецифических экспериментальных аутоиммунных заболеваний у животных.
Дифференцировка и регуляция CD4+ Т-клеток зависят от множества факторов, включая силу взаимодействия антиген-антитело, степень костимуляции, цитокины, присутствующие в среде, экспрессию факторов транскрипции и их взаимодействие с модификаторами гистонов. Во время развития тимоцитов костимуляция CD154 играет важную роль в разнообразии репертуара TCR. Дефицит CD154 ослабляет совместное использование клона TCRβ по сравнению с клоном дикого типа в зависимых от Т-клеток иммунных реакциях, что приводит к неправильному редактированию клонотипов Т-клеток во время отрицательной селекции тимуса (Fähnrich et al.). По мере того, как оценка пластичности подмножества Т-хелперов возрастает, их характеристика становится еще более важной. Отчетливо противоположные субпопуляции Т-хелперов могут экспрессировать один и тот же рецептор, секретировать общий цитокин и регулироваться одним и тем же транскрипционным фактором. Исследование Huang et al. призывает к осторожности при использовании LAG3/CD49. b коэкспрессия в качестве автономных маркеров для идентификации клеток Tr1, поскольку они также могут экспрессироваться T-клетками Foxp3+ и CD8+ T-клетками. Дальнейшие исследования позволят проанализировать физиологическую значимость экспрессии этих маркеров в различных подмножествах Т-клеток. Несколько исследований показали участие различных субпопуляций Т-хелперов в патофизиологии воспалительных заболеваний (4). В модели нейровоспаления было продемонстрировано, что клетки Th27 получают помощь от клеток Tfh для воспалительного В-клеточного ответа (Quinn et al.). В-клетки, регулируемые клетками Tfh, могут перемещаться в ЦНС и подвергаться переключению классов, что коррелирует с тяжестью заболевания (Quinn et al.).
b коэкспрессия в качестве автономных маркеров для идентификации клеток Tr1, поскольку они также могут экспрессироваться T-клетками Foxp3+ и CD8+ T-клетками. Дальнейшие исследования позволят проанализировать физиологическую значимость экспрессии этих маркеров в различных подмножествах Т-клеток. Несколько исследований показали участие различных субпопуляций Т-хелперов в патофизиологии воспалительных заболеваний (4). В модели нейровоспаления было продемонстрировано, что клетки Th27 получают помощь от клеток Tfh для воспалительного В-клеточного ответа (Quinn et al.). В-клетки, регулируемые клетками Tfh, могут перемещаться в ЦНС и подвергаться переключению классов, что коррелирует с тяжестью заболевания (Quinn et al.).
Исследователи накопили значительный объем информации о влиянии CD4+ T-клеток на различные воспалительные заболевания у мышей и людей. Поведение CD4+ Т-клеток регулируется внутренними метаболическими свойствами. Метаболиты липидов могут действовать как регуляторы иммунных реакций.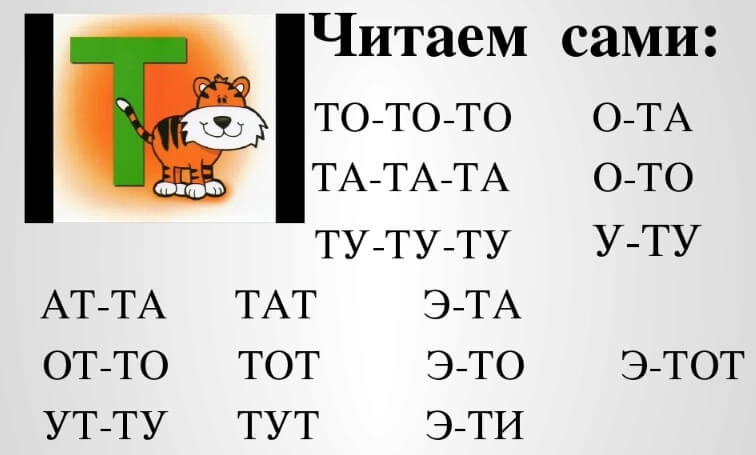 Изменение путей поступления стероидов может влиять на воспаление и отвечать за патофизиологию различных заболеваний. Фермент холестерин-25-гидроксилаза, который синтезирует 25-OHC, может усиливать индуцированный IL-27 T 9.0085 r 1 клетки (Vigne et al.). 25-OHC может отрицательно регулировать продукцию IL-10 клетками Tr1. Метаболизм Т-клеток был нацелен на эффективную иммунотерапию рака и изменение функций эффекторных Т-клеток [(5), Roy et al.]. Для активации и пролиферации Т-клеток глюкоза обеспечивает необходимую энергию (Roy et al.). Дополнительные метаболиты, включая липиды, АТФ, оксид азота, НАД, также играют решающую роль в дифференцировке CD4+ Т-клеток. И mTOR, и AMPK являются сенсорами, которые регулируют метаболические контрольные точки дифференцировки Т-клеток. Ингибиторы mTOR могут ослаблять гликолиз, вызывая дифференцировку Т-клеток памяти, в то время как ингибиторы AMPK снижают метаболизм жирных кислот, что, в свою очередь, способствует дифференцировке клеток Th2 и Th27.
Изменение путей поступления стероидов может влиять на воспаление и отвечать за патофизиологию различных заболеваний. Фермент холестерин-25-гидроксилаза, который синтезирует 25-OHC, может усиливать индуцированный IL-27 T 9.0085 r 1 клетки (Vigne et al.). 25-OHC может отрицательно регулировать продукцию IL-10 клетками Tr1. Метаболизм Т-клеток был нацелен на эффективную иммунотерапию рака и изменение функций эффекторных Т-клеток [(5), Roy et al.]. Для активации и пролиферации Т-клеток глюкоза обеспечивает необходимую энергию (Roy et al.). Дополнительные метаболиты, включая липиды, АТФ, оксид азота, НАД, также играют решающую роль в дифференцировке CD4+ Т-клеток. И mTOR, и AMPK являются сенсорами, которые регулируют метаболические контрольные точки дифференцировки Т-клеток. Ингибиторы mTOR могут ослаблять гликолиз, вызывая дифференцировку Т-клеток памяти, в то время как ингибиторы AMPK снижают метаболизм жирных кислот, что, в свою очередь, способствует дифференцировке клеток Th2 и Th27. Напротив, активация пути AMPK может оказывать обезболивающее действие при воспалительной боли путем ослабления Il1 экспрессия β и блокирование активации NF-κB (6). В наивных Т-клетках наблюдается повышенное отношение АМФ к АТФ в отсутствие передачи сигналов TCR, что приводит к уменьшению mTOR и устойчивой функции AMPK (7). Интересно, что во время лаг-фазы активированных Т-клеток индуцированные цитозольные ионы кальция способствуют функции AMPK, несмотря на снижение уровня AMP. Однако в фазе роста активированных Т-клеток повышение уровня АТФ приводит к устойчивой функции mTOR. Истощение аминокислот Trp и Arg может ослабить как активацию, так и функцию эффекторных Т-клеток (5). Снижение уровня кислорода и окислительного фосфорилирования может увеличить экспрессию PD-L1 на раковых клетках. Активация HIF-1 может индуцировать эффекторные свойства Т-клеток за счет усиления гликолиза и глутаминолиза. Другие факторы транскрипции, которые могут действовать как метаболические контрольные точки во время дифференцировки Т-клеток, включают BCL-6 (дифференцировка клеток Tfh), IRF4 (Th3, Th27, Th9дифференцировка клеток), Foxo (дифференциация клеток Th9), MYC (баланс между дифференцировкой клеток Th27 и Treg).
Напротив, активация пути AMPK может оказывать обезболивающее действие при воспалительной боли путем ослабления Il1 экспрессия β и блокирование активации NF-κB (6). В наивных Т-клетках наблюдается повышенное отношение АМФ к АТФ в отсутствие передачи сигналов TCR, что приводит к уменьшению mTOR и устойчивой функции AMPK (7). Интересно, что во время лаг-фазы активированных Т-клеток индуцированные цитозольные ионы кальция способствуют функции AMPK, несмотря на снижение уровня AMP. Однако в фазе роста активированных Т-клеток повышение уровня АТФ приводит к устойчивой функции mTOR. Истощение аминокислот Trp и Arg может ослабить как активацию, так и функцию эффекторных Т-клеток (5). Снижение уровня кислорода и окислительного фосфорилирования может увеличить экспрессию PD-L1 на раковых клетках. Активация HIF-1 может индуцировать эффекторные свойства Т-клеток за счет усиления гликолиза и глутаминолиза. Другие факторы транскрипции, которые могут действовать как метаболические контрольные точки во время дифференцировки Т-клеток, включают BCL-6 (дифференцировка клеток Tfh), IRF4 (Th3, Th27, Th9дифференцировка клеток), Foxo (дифференциация клеток Th9), MYC (баланс между дифференцировкой клеток Th27 и Treg). Были разработаны лекарства, воздействующие на эти различные метаболические контрольные точки для облегчения различных воспалительных заболеваний, включая болезнь Крона, язвенный колит, диабет 2 типа, ревматоидный артрит и хроническую обструктивную болезнь легких. Активация Т-клеток также опосредована незаменимыми микроэлементами, включая цинк. Стадии передачи сигналов TCR потенциально могут быть изменены цинком (8). Транспортер цинка Zip6, экспрессируемый на поверхности нестимулированных Т-клеток, рекламируется как важный, снижающий порог активации Т-клеток. Витамины также регулируют активацию Т-клеток. Несмотря на то, что витамин D блокирует пролиферацию CD4+ T-клеток, он увеличивает количество CD4+CD25+Foxp3+ Treg-клеток. Следовательно, нацеливание на Т-клетки с помощью регуляторов метаболизма представляет собой захватывающее направление для дальнейших исследований по регулированию патофизиологии воспалительных заболеваний и рака.
Были разработаны лекарства, воздействующие на эти различные метаболические контрольные точки для облегчения различных воспалительных заболеваний, включая болезнь Крона, язвенный колит, диабет 2 типа, ревматоидный артрит и хроническую обструктивную болезнь легких. Активация Т-клеток также опосредована незаменимыми микроэлементами, включая цинк. Стадии передачи сигналов TCR потенциально могут быть изменены цинком (8). Транспортер цинка Zip6, экспрессируемый на поверхности нестимулированных Т-клеток, рекламируется как важный, снижающий порог активации Т-клеток. Витамины также регулируют активацию Т-клеток. Несмотря на то, что витамин D блокирует пролиферацию CD4+ T-клеток, он увеличивает количество CD4+CD25+Foxp3+ Treg-клеток. Следовательно, нацеливание на Т-клетки с помощью регуляторов метаболизма представляет собой захватывающее направление для дальнейших исследований по регулированию патофизиологии воспалительных заболеваний и рака.
В совокупности статьи, опубликованные в рамках темы исследования, освещают новые роли и основные механизмы дифференцировки и функций Т-клеток при воспалении тканей и их влияние на патогенез воспалительных заболеваний. Основываясь на опубликованной работе по этой теме, необходимо также понять функциональную динамику пластичности Th-клеток, которая приводит к конечному результату иммунного патогенеза заболеваний и приводит к продвижению нашего понимания иммунологических основ заболеваний. Полученные иммунологические знания из опубликованных статей будут способствовать дальнейшему совершенствованию и новым иммунным стратегиям лечения воспалительных состояний.
Основываясь на опубликованной работе по этой теме, необходимо также понять функциональную динамику пластичности Th-клеток, которая приводит к конечному результату иммунного патогенеза заболеваний и приводит к продвижению нашего понимания иммунологических основ заболеваний. Полученные иммунологические знания из опубликованных статей будут способствовать дальнейшему совершенствованию и новым иммунным стратегиям лечения воспалительных состояний.
Вклад авторов
RG и AA задумали, разработали и написали рукопись. Все авторы прочитали и одобрили окончательный вариант рукописи для публикации.
Финансирование
Эта работа была поддержана средствами Департамента биотехнологии и Департамента науки и техники правительства Индии для AA. RG была поддержана грантом INNO-INDIGO, Департамента науки и технологий правительства Индии.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.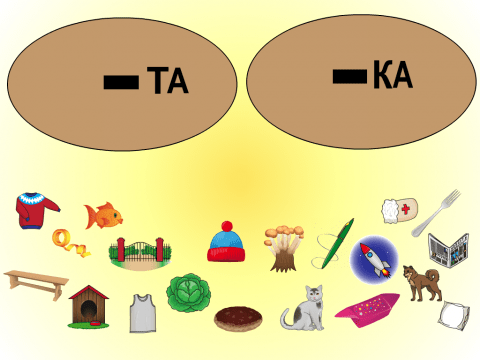
Благодарности
Мы признательны и признательны всем авторам и их участие в этой теме исследования. Мы выражаем благодарность всем рецензентам за согласие участвовать в процессе рецензирования и предоставление своих комментариев и отзывов о рукописи.
Ссылки
1. Хаскинс К., Кубо Р., Уайт Дж., Пиджен М., Капплер Дж., Маррак П. Антигенный рецептор, ограниченный главным комплексом гистосовместимости, на Т-клетках. I. Выделение моноклональными антителами. J Exp Med. (1983) 157:1149–69. doi: 10.1084/jem.157.4.1149
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
2. Хашимото К., Коно Т., Икава Т., Хаяцу Н., Миядзима Ю., Ябуками Х. и др. Транскриптомика одиночных клеток выявляет экспансию цитотоксических Т-клеток CD4 у долгожителей. Proc Natl Acad Sci USA. (2019) 116:24242–51. doi: 10.1073/pnas.1907883116
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
3. Мосманн Т.Р.
Реферат PubMed | Google Scholar
4. Chemin K, Gerstner C, Malmstrom V. Эффекторные функции CD4+ T-клеток в месте локального аутоиммунного воспаления – уроки ревматоидного артрита. Фронт Иммунол. (2019) 10:353. doi: 10.3389/fimmu.2019.00353
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
5. Le Bourgeois T, Strauss L, Aksoylar HI, Daneshmandi S, Seth P, Patsoukis N, et al. Нацеливание на метаболизм Т-клеток для улучшения иммунотерапии рака. Фронт Онкол. (2018) 8:237. doi: 10.3389/fonc.2018.00237
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
6. Xiang HC, Lin LX, Hu XF, Zhu H, Li HP, Zhang RY, et al. Активация AMPK ослабляет воспалительную боль за счет ингибирования активации NF-kappaB и экспрессии IL-1beta.